地磁场无处不在,为许多动物提供了导航的“指南针”。过去五十多年的行为学研究已揭示:从迁徙的候鸟到归家的信鸽,许多动物都能感知并利用地磁场进行定向和导航。然而,动物磁感知的神经机制仍是未解之谜。目前有三种主流的磁感知机制假说:磁铁矿介导的磁感知假说(Winklhofer et al., 2010)、光依赖性自由基对磁感知假说(Mouritsen et al., 2012)、电压门控通道介导的电磁感应假说(Nimpf et al., 2019),这些假说分别与三叉神经、视网膜、内耳等解剖结构相关。然而,以往的研究大多局限于对特定脑区的检验,可能带有先入为主的偏见。要真正理解磁感知机制,需要一种不受既有理论束缚的全局性视角。
为了打破这一局限,德国慕尼黑大学的David Keays教授领导的研究团队采用组织透明化技术,结合绘制全脑神经元活动图谱的全新策略,尝试回答一个核心问题:在完整的鸽子大脑中,究竟哪些神经元群体(脑区)会对磁场刺激产生响应?通过精准定位这些被激活的“核心脑区”,他们逐步构建出感磁神经回路,再逆向追踪至外周,最终锁定了初始的磁感知受体(Nordmann et al., 2026)。这一系统性的研究路径,首次在鸽子大脑中明确了与磁感知相关的脑区和神经回路,为理解鸟类为何具备非凡的导航能力,提供了关键证据。
建立全脑活动图谱,筛选核心磁激活脑区
为了建立全脑活动图谱并筛选出核心的磁激活脑区,研究人员首先对鸽子大脑的组织透明化技术进行了优化。经过处理后,整个鸽子的大脑变得透明,可以在光片显微镜下实现全脑三维成像,结合C-FOS标记就可精准定位被激活神经元的具体位置和数量。在此基础上,团队设计了多组对照实验。他们在有光和无光条件下,分别让鸽子暴露于旋转磁场或对照零磁场环境中,然后将受磁场刺激组的大脑活动图谱与对照组进行比对。结果发现两个核心激活区域:一个是尾侧内侧前庭核(VeM),该区域呈双侧对称激活,主要负责接收前庭系统的传入信号,提示前庭系统可能参与了磁感知信息的处理,也暗示着磁信号的初级感受器可能位于前庭上皮。另一个是尾侧中脑皮层(MC),这是一个与多感官整合相关的脑区,其激活区域沿海马脑室分布,位于听觉区L2的内侧。即使在无光黑暗的条件下,磁刺激依然能持续激活前庭核;而视觉相关脑区(如背外侧膝状体复合体、视顶盖、视觉 Wulst)则未见显著激活,这表明其潜在机制并不依赖光诱导自由基对的形成。
研究团队同时还设计了静磁场对照实验。结果显示:暴露于静磁场的鸽子,其内侧前庭核、中脑皮层、背内侧丘脑、海马均未被显著激活。这一结果证实,变化的磁场才是激活这些脑区的必要条件,这与依赖于电磁感应的生物物理机制相一致。
向内耳深入:寻找感受磁场的细胞
全脑筛选结果将研究焦点指向了前庭系统。顺着这条线索,研究人员进一步深入探索前庭信号的发源地——内耳的壶腹嵴。壶腹嵴位于各半规管的基底,是感受头部旋转的关键结构,其中分布着感觉毛细胞。电磁感应假说能否成立,有一个核心前提:感受细胞中必须存在能够感知电信号的分子装置。那么,鸽子的壶腹嵴是否具备这样的分子基础?为了回答这个问题,研究者对壶腹嵴细胞进行了单细胞RNA测序。分析结果显示,一类II型毛细胞中高表达两种关键的离子通道——BK钾通道和钙通道CaV1.3异构体。此前已有研究表明,CaV1.3异构体是电感知的关键分子(Bellono et al., 2018)。这一发现为电磁感应机制提供了重要的分子证据。综合这些结果,II型毛细胞很可能是鸽子感知磁场的初级感受细胞。通过CaV1.3与BK通道的协同作用,这些细胞具备了将磁场诱导的微弱电压变化转换为细胞电信号的分子能力。
构建基于内耳电磁感应的磁感知回路模型
综合全脑活动图谱与单细胞分子图谱,研究团队提出了一套完整的磁感知模型(图1)。该模型的起点是物理信号的出现:当鸽子头部运动方向与地磁场方向垂直时,根据电磁感应原理,半规管内的内淋巴液中会诱导出微弱的电场。这一物理信号随即被细胞层面的分子装置捕获——II型毛细胞通过其表面的CaV1.3钙通道和BK钾通道感知电压变化,将物理磁场信号转换为细胞可读的生物电信号。整个过程不依赖光的参与,与光诱导自由基对机制形成鲜明对比。电信号随后沿着神经通路传递:从前庭系统的壶腹嵴出发,经由前庭耳蜗神经进入脑干,首先抵达内侧前庭核进行初步处理;继而通过背内侧丘脑的中继,最终上传至两个高级脑区——尾侧中脑皮层和海马体。这两个区域分别负责多感官整合与空间导航记忆,共同构成了鸽子感知地磁场并实现精准定位导航的神经基础。
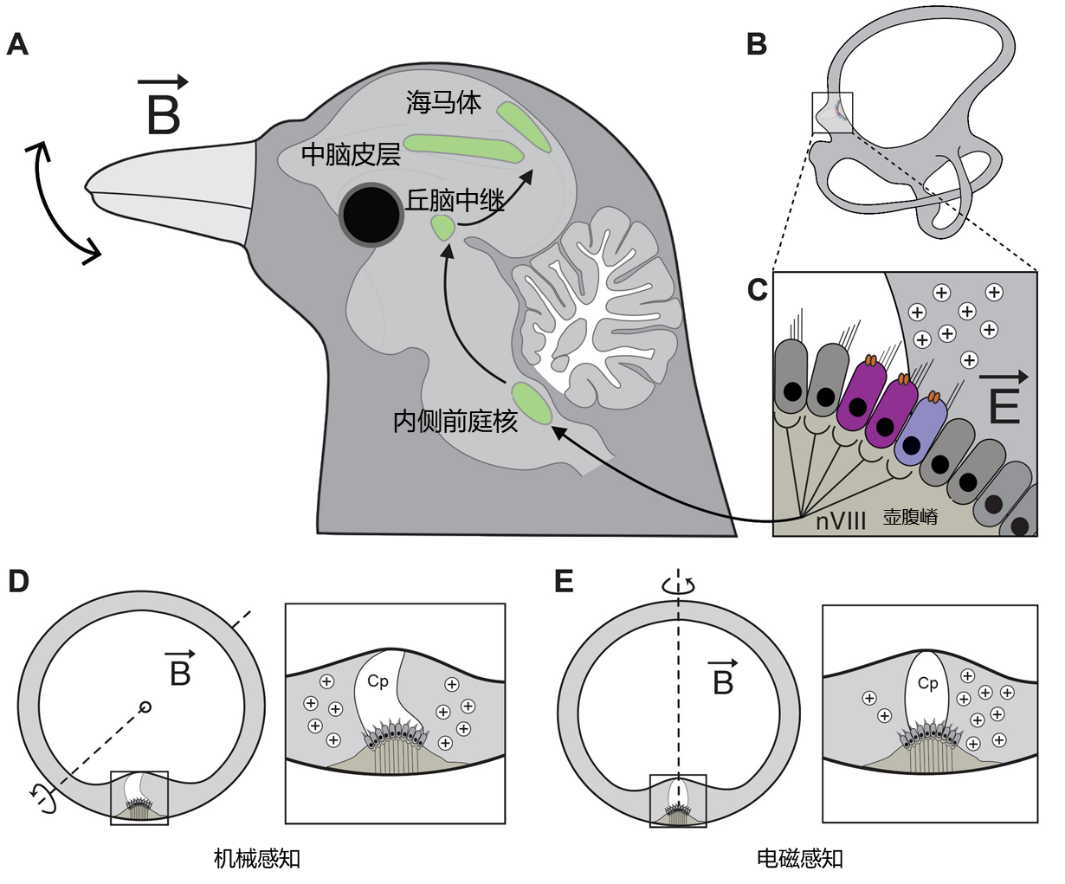
图1 电磁感应驱动的磁感知回路模型。 (A至C) 头部运动产生的变化磁场在半规管中诱导出电场(E),导致内淋巴液内形成电压梯度(B)。被凝胶状的壶腹帽分隔开的这种电荷差异,由表达电压敏感性钙通道CaV1.3和大电导钙激活钾通道BK的II型毛细胞检测(C)。这一电磁信息从壶腹嵴(ca)经由前庭耳蜗神经(nVIII)传至内侧前庭核(VeM)进行初步处理,随后通过丘脑中继(DTh),与中脑皮层(MC)和海马体(Hp)内的其他感觉信息整合。(D) 头部在半规管平面内的旋转通过壶腹帽(Cp)的位移导致毛细胞静纤毛的机械性刺激,不产生电磁感应。(E) 相反,在存在磁场向量(B)的情况下,头部垂直于半规管平面的旋转会导致电荷在壶腹帽两侧重新分布,而毛细胞不发生机械位移。因此,鸟类能够区分机械性输入和电磁性输入
意义与展望
这项研究的意义在于:它首次以全局、无偏见的视角,系统地描绘了鸽子大脑对磁场刺激的响应图谱,为“电磁感应假说”提供了迄今为止最有力的神经解剖学和分子生物学证据。它成功地将磁场感知的起点指向了内耳前庭系统,并鉴定出了潜在的“感应分子”——CaV1.3和BK通道。研究还排除了其它两类磁感知机制的可能性。
然而,这项突破也开启了更多待解之谜。正如作者所言,一个持续的挑战是证明这些特定分子和细胞类型对于磁感知的“必要性和充分性”。未来的研究需要借助基因编辑、病毒示踪和电生理记录等更精密的技术进行验证,例如特异性敲除II型毛细胞中的CaV1.3通道,观察鸽子是否因此失去磁感应能力。此外,中脑皮层和海马体中的神经元如何编码磁倾角、极性和强度等具体磁参数,以及这一系统如何与其他感官信息整合形成完整的导航能力,这些都是未来研究需要回答的重要问题。尽管挑战犹存,这项研究无疑为我们最终揭开动物“第六感”的神秘面纱迈出了关键一步。
主要参考文献
Bellono N W, Leitch D B, Julius D. Molecular tuning of electroreception in sharks and skates[J]. Nature, 2018, 558(7708): 122-126.
Mouritsen H, Hore P J. The magnetic retina: light-dependent and trigeminal magnetoreception in migratory birds[J]. Current opinion in neurobiology, 2012, 22(2): 343-352.
Nimpf S, Nordmann G C, Kagerbauer D, et al. A putative mechanism for magnetoreception by electromagnetic induction in the pigeon inner ear[J]. Current Biology, 2019, 29(23): 4052-4059. e4.
Nordmann G C, Balay S D, Kapuruge T N, et al. A global screen for magnetically induced neuronal activity in the pigeon brain[J]. Science, 2025: eaea6425.(原文链接)
Winklhofer M, Kirschvink J L. A quantitative assessment of torque-transducer models for magnetoreception[J]. Journal of the Royal Society, Interface, 2010, 7 Suppl 2(Suppl 2): S273-S289.
(撰稿:田兰香、赵晨晨/地球与行星物理学科中心)